A Refined Kinetic Analysis of Plasminogen Activation by Recombinant Bovine
Tissue-Type Plasminogen Activator Indicates Two Interconvertible Activator
Forms
†
Laust B. Johnsen,
§
Peter Ravn,
§,⊥
Lars Berglund,
§
Torben E. Petersen,*
,§
Lone K. Rasmussen,
§
Christian W. Heegaard,
§
Jan T. Rasmussen,
§
Connie Benfeldt,
|
and Sergey N. Fedosov
§
Protein Chemistry Laboratory, Department of Molecular and Structural Biology, UniVersity of Aarhus, Science Park,
GustaV Wieds Vej 10 C, DK-8000 Aarhus C, Denmark, and MD Foods Research and DeVelopment Centre, RørdrumVej 2,
DK-8220 Brabrand, Denmark
ReceiVed March 23, 1998; ReVised Manuscript ReceiVed June 18, 1998
ABSTRACT: Bovine tissue-type plasminogen activator (tPA) was heterologously expressed in the methylo-
trophic yeast Pichia pastoris and characterized structurally and kinetically. The bovine single-chain tPA-
mediated activation of bovine plasminogen was studied in the presence and absence of fibrinogen fragments.
We have proposed a refined new method of kinetic analysis which allows examination of both stationary
and prestationary phases of this process. The investigation revealed the presence of two interconvertible
forms of the recombinant bovine tPA being in equilibrium ata1to50ratio. Only the minor form was
able to bind and activate plasminogen. Saturation of the whole pool of tPA required high plasminogen
concentration (K
m
g 5µM) in order to reverse the equilibrium between the two forms. Fibrinogen fragments
activated the single-chain tPA due to preferential binding and stabilization of the minor “active” form of
the enzyme until all the molecules of tPA were converted. The same mechanism could be applied to
human tPA as well. The K
m
values, obtained for recombinant bovine and human tPA in the presence of
fibrinogen fragments, were found to be similar (K
m
) 0.1 µM) while k
cat
of human tPA was 5-10 times
higher.
The activation of plasminogen is an important process
associated with degradation of an extracellular matrix such
as dissolving of blood clots, tissue remodeling, and invasive
growth of cancer cells. Tissue-type plasminogen activator
(tPA)
1
and urokinase-type plasminogen activator (uPA) are
the two major proteins responsible for conversion of plas-
minogen to plasmin. Although both tPA and uPA are related
enzymes and activate plasminogen by cleavage of the same
peptide bond, they have their own physiological features.
The tPA-induced process is stimulated significantly on the
surface of fibrin, and tPA is regarded as the fibrinolytic
activator. On the other hand, the pericellular uPA-mediated
activation of plasminogen is supposed to be engaged in tissue
remodeling and cancer metastasis development.
The activation of plasminogen has been studied in detail
in the human system from where the involved protein
components have been identified and characterized. In
contrast much less is known about plasminogen activation
in other species. Bovine mastitis is an inflammatory disease
of the mammary gland induced by various microorganisms,
and a 20-fold increase in tPA activity has been reported in
the milk of cows infected with Staphylococcus aureus (1).
The activation of plasminogen by tPA is greatly increased
by the R
s2
-casein dimer (2), and in order to study this system
in more detail it is necessary to obtain bovine tPA. As this
protein is only present in very small amounts in natural
tissues and fluids and to our knowledge never has been
purified, we have made recombinant expression of the
protein.
tPA is a mosaic protein with five domains consisting of a
finger domain, an epidermal growth factor domain, two
kringle domains, and a serine proteinase domain. The finger
and the second kringle are believed to be responsible for
the interaction with fibrin (3-5). tPA is synthesized as a
single-chain polypeptide and can be converted into its more
active two-chain form by plasmin scission of an Arg-Ile bond
situated in the strand connecting kringle 2 with the serine
proteinase domain (6).
Kinetic analysis of the human tPA catalysis was rendered
in a number of papers (7-9) and resulted in several
conclusions concerning its mechanism. The single-chain
enzyme was considered as a poor catalyst when compared
to the double-chain form. It was characterized by K
m
) 1-2
µM toward [Glu
1
]plasminogen and k
cat
) 0.3-0.6 min
-1
(9).
†
This work is part of the FØTEK program supported by the Danish
Dairy Research Foundation (Danish Dairy Board) and the Danish
Government.
* Corresponding author.
§
University of Aarhus.
|
MD Foods Research and Development Centre.
⊥
Present address: Biotechnological Institute, Kogle Alle´ 2, DK-
2970 Hørsholm, Denmark.
1
Abbreviations: [Asp
1
]plasminogen, native form of bovine plas-
minogen with an aspartic acid at the N-terminus; EACA, -aminocaproic
acid; Fb, fibrinogen; [Glu
1
]plasminogen, native form of human plas-
minogen with a glutamic acid at the N-terminus; PAI, plasminogen
activator inhibitor; Pg, plasminogen; Pn, plasmin; tPA, tissue-type
plasminogen activator; uPA, urokinase plasminogen activator.
12631Biochemistry 1998, 37, 12631-12639
S0006-2960(98)00669-2 CCC: $15.00 © 1998 American Chemical Society
Published on Web 08/19/1998

At the same time, the double-chain tPA had a considerably
higher affinity to the substrate K
m
) 0.1-0.5 µM as well as
higher ability for plasminogen transformation k
cat
) 4-5
min
-1
(8). All catalytic characteristics of the single-chain
tPA improved when fibrin (or fibrinogen fragments) was
added to the medium, showing the K
m
and k
cat
values at the
same level as those of the double-chain enzyme (7, 9). The
double-chain tPA also demonstrated sensitivity to fibrin
which was manifested in 5-10-fold increase of k
cat
but
without any changes in K
m
(8). The existence of an active
ternary complex plasminogen-fibrin-tPA, converting bound
plasminogen to plasmin, was proposed to be more favorable
than the action of a fibrin-tPA complex toward free
plasminogen (9). Previous publications about tPA kinetics
mainly concerned the tPA-catalyzed reaction being in steady
state. This is, in part, due to mathematical difficulties in
the approximation of the complex reaction when the tPA-
catalyzed conversion of plasminogen to plasmin was fol-
lowed by cleavage of a measurable substrate by plasmin.
The human tPA might be an inconvenient object for such
investigations as well. Therefore, application of tPA from
another source, provided with an adapted mathematical
mechanism, could shed some light on “hidden” stages of
the process. An appropriate candidate for this inquiry is
bovine tPA, promising to be useful in several aspects.
We have expressed bovine tPA in the methylotrophic yeast
Pichia pastoris and characterized the product from a
structural and kinetical point of view. Comparison between
human and bovine tPA required standardization of the
experimental conditions, therefore activation of both enzymes
was induced by bovine fibrinogen fragments. The bovine
tPA was characterized by a slower equilibration with the
components of the reaction medium when compared to
human tPA. This allowed a detailed analysis of prestationary
kinetics, impossible under the same conditions for the
catalytic reaction performed with human tPA.
MATERIALS AND METHODS
Chemicals and Reagents. P. pastoris GS115 (his4) (10),
the protease deficient strain SMD1168 (his4, pep4), and the
expression vectors pPIC9K and pHIL-D2 were purchased
from Invitrogen Corp. Super Taq polymerase was from HT
Biotechnology, nucleotide triphosphates were from Pharma-
cia, Chameleon mutagenesis kit was from Stratagene, oligo-
nucleotides were from DNA technology (Science park,
Aarhus, Denmark), and all other enzymes were from New
England Biolabs. PCR was performed in a Hybaid ABA-
CUS thermal cycler. Sequencing was performed either with
a Sequenase kit version 2.0 from United States Biochemical
Corporation or with a dye terminator cycle sequencing kit
from Perkin-Elmer. [
35
S]dATP was from Amersham Inter-
national. Sequencing, ligation, transformation of Escherichia
coli, DNA preparation, PCR, and other DNA modifying
processes were performed according to the manufacturers’
recommendations or standard laboratory procedures. Yeast
media were composed as follows: YPD (1% yeast extract,
2% peptone, 2% dextrose), BMGY (1% yeast extract, 2%
peptone, 100 mM potassium phosphate pH 6.0, 1.34% YNB,
1% glycerol), BMMY (1% yeast extract, 2% peptone, 100
mM potassium phosphate pH 6.0, 1.34% YNB, 1% metha-
nol), MM (1.34% YNB, 0.00004% biotin, 1% methanol),
and MD (1.34% YNB, 0.00004% biotin, 1% dextrose).
Yeast extract, peptone, YNB, and casamino acids were from
DIFCO, and G418 was from Life Technologies. Recombi-
nant human tPA (Actilyse) and chromozym tPA (CH
3
-SO
2
-
D-Phe-Gly-Arg-pNA) were from Boehringer Mannheim.
S-2251 (H-
D-Val-Leu-Lys-pNA) was from Chromogenix.
Bovine [Asp
1
]plasminogen and PAI-1 were from American
Diagnostica. PAI-2 was of the low molecular weight form
and generously provided by Dr. I. Lecander (Lund, Sweden).
CNBr-fibrinogen fragments were made by incubation of 100
mg fibrinogen with 130 mg CNBr in 70% formic acid at
room temperature for 16 h. The resulting fibrinogen
fragments were dialyzed against water for removal of low
molecular weight fragments (membrane cut off ) 12-14
kDa) and stored at -80 °C at a concentration of 2.8 mg/
mL. The bovine plasminogen used for zymography was
purified as described in ref 11, and plasminogen depleted
bovine fibrinogen was obtained from Enzyme Research
Laboratories.
Construction of the P. pastoris tPA Expression Vectors.
pHIL-D2/tPA was derived from the P. pastoris integration
vector pHIL-D2. The tPA encoding region, including the
native signal peptide, was inserted into the EcoRI restriction
site in pHIL-D2, yielding pHIL-D2/tPA. Due to the presence
of an internal EcoRI restriction site in the tPA cDNA, this
was performed by PCR on pBtPA4 (12) with the forward
5′-ATGATGAGCGCAATGAAG-3′ and reverse 5′-GGT-
GTCCCTGGTCATGG-3′ primers. The EcoRI restriction
site in the resulting amplified tPA encoding region was then
methylated by EcoRI methylase and ligated to EcoRI linker
oligonucleotides 5′-GGAATTCC-3′. Following digestion by
EcoRI, the amplified fragment was ligated into pHIL-D2.
pPIC9K/tPA was derived from the P. pastoris integration
vector pPIC9K. Using PCR, 5′ and 3′ NotI restriction sites
were introduced into the bovine tPA cDNA using the plasmid
pBtPA4 as template and the following forward and reverse
primers, respectively, 5′-CTCAGGAGAGCGGCCGCATCG-
TAC-3′ and 5′-GAGGAAAGCGGGCGGCCGCCCTGGG-
3′. The resulting PCR product was cloned into the NotI
restriction site in pPIC9K. To generate the native N-terminus
of tPA after cleavage by the signal peptidases (Figure 1),
site specific mutagenesis was performed on pPIC9K/tPA,
using the primer 5′-CTCGAGAAAAGAGAGGCTGAAGCT-
TCGTACAAAGTGACCTGCAGAGAT-3′. To ensure that
the T4 polymerase replicated the entire plasmid, two non-
mutagenic primers, situated in the Col E1 region (7961-
7980) and HIS4 region (4801-4821) of pPIC9K/tPA, were
included in the mutagenesis reaction. The tPA encoding
region of pPIC9K/tPA was finally sequenced as a control
for PCR-introduced mutations.
Transformation of P. pastoris with pPIC9K/tPA and pHIL-
D2/tPA and Multicopy Colony Selection. P. pastoris strains
GS115 (his4) and SMD1168 (his4, pep4) were transformed
with the expression plasmids using the spheroblasting and
FIGURE 1: Partial amino acid sequence of the pPIC9K/tPA secretion
signal fused to the mature N-terminal of tPA is shown, as well as
the expected two endoproteolytic split sites (KEX2, STE13).
12632 Biochemistry, Vol. 37, No. 36, 1998 Johnsen et al.

electroporation techniques. Mut
+
and Mut
s
phenotypes were
determined by screening for fast and slow growth on
methanol-containing plates (MM plates) and, as a control,
evaluating the growth of the same colonies on dextrose-
containing plates (MD plates). Multicopy colonies were
selected by their ability to grow on YPD plates containing
increasing concentrations of G418 (0.25-4 mg/mL). The
transformation, phenotype determination, and selection were
done essentially as described by the supplier.
Fermentation of pPIC9K/tPA Transformed P. pastoris
Strains. Cells were restreaked from freeze cultures on YPD
or MD plates and a single colony was used for inoculation
of 10 mL of BMGY medium. Cells were grown to log
phase, and a suitable volume was used to inoculate 100 mL
of BMGY medium, after which cells were grown until an
OD
600
of 2-6 was reached. Subsequently, the cells were
centrifuged at 1000g at room temperature, the BMGY
medium was discarded, and the cells were resuspended into
200 mL of BMMY medium (with or without 1% casamino
acids) in 2 L baffled shake flasks to an OD
600
) 1. Cells
were grown aerobically (shake flask covered with gaze) and
supplemented with 1% methanol every 24 h. Cells were
removed by centrifugation, and the supernatant was stored
at -80 °C. Cells were always grown in a rotary shaker at
30 °C, 300 rpm. When the expression levels of different
strains were compared, fermentation was performed in a 12
mL reagent glass essentially as described above.
Purification of tPA. All steps were carried out at 4 °C.
Two hundred milliliters of the supernatant was thawed,
centrifuged (10 min, 10000g), dialyzed against 20 mM NaH
2
-
PO
4
, pH 7.0, 100 mM NaCl, 0.05% Tween 80, and applied
to a column of 40 mL of lysine-Sepharose (prepared by
reaction of CNBr-activated Sepharose 4B with lysine). After
the column was loaded, it was washed with 400 mL of 20
mM NaH
2
PO
4
, 200 mM NaCl, and 0.05% Tween 80 and
eluted with the same buffer containing 200 mM
L-arginine.
Fractions were screened for tPA activity, pooled, and
concentrated on an Amicon-30 membrane until A
280
was
approximately 1. The purified protein was stored at -80
°C for later use.
tPA ActiVity Assay and Zymography. tPA activity in
international units (U) was determined by use of the
chromogenic substrate chromozym t-PA according to the
assay described by the manufacturer. The assay was
performed in a total volume of 0.2 mL at 37 °C in microtiter
plates by the addition of the tPA sample to 0.1 M Tris pH
8.5, 0.15% Tween 80, and 0.4 mM chromozym t-PA. The
reaction was followed at 405 nm over a period of1hina
thermostatically controlled Bio-Tek EL 340 BioKinetics
Reader (Bio-Tek Instruments Inc., Winooski, VT). The
amount of U/mL was calculated as follows: (measured
absorbance × min
-1
× mL
-1
tPA sample) × (1 cm/ 0.6 cm,
correction for light path in microtiter wells) × (1/9.75, U
conversion factor). The zymography was performed as
described in ref 13.
Coupled Peptidyl Anilide Plasminogen ActiVation Assay.
The plasminogen activation potential of tPA was evaluated
in a coupled peptidyl anilide assay, where the formation of
plasmin was measured by its hydrolysis of the chromogenic
substrate S-2251. The plasminogen activation reaction was
performed in a total of 0.2 mL containing 0.1 M Tris pH
7.4, 0.02% Tween 80, 0.03-1.08 µM plasminogen, (8.4
µg fibrinogen fragments, and 0.5 mM S-2251. The reaction
was initiated by addition of bovine tPA to the final
concentration of 0.01 µM in the absence of fibrinogen
fragments or 0.001 µM tPA in their presence. The concen-
tration of human tPA was 3.3 × 10
-4
µM in both cases.
The reactions were carried out at 37 °C in microtiter plates
and were followed at 405 nm over a period of1hinthe
same spectrometer as the tPA activity assay. For each assay,
at least two independent experiments were made, with double
determination in each experiment.
Mass Spectroscopy and Amino Acid Sequence Analysis.
Automated Edman degradation was carried out on an ABI
477A/120A protein sequencer (Applied Biosystems) using
standard programs. Mass spectra were acquired using a
matrix-assisted laser desorption ionization (MALDI) mass
spectrometer (Bruker BIFLEX, Bruker-Franzen, Bremen,
Germany) equipped with a nitrogen ultraviolet laser at 337
nm. Samples (2 µL) dissolved in 0.1% trifluoroacetic acid
were mixed with 2 µL R-cyano-4-hydroxycinnamic acid (15
g/L).
Kinetic Analysis of the Coupled Peptidyl Anilide Plasmi-
nogen ActiVation Assay. Three kinetic models were designed
in order to fit the experimental data. The basic model 1
implies the existence of two reactions: (i) transformation
of plasminogen to plasmin catalyzed by tPA, and (ii)
utilization of a chromogenic substrate by plasmin. A new
method of linearization of the initial coordinates (product
versus time) is proposed. A more complex scheme, de-
scribed in model 2, considers the existence of a prestationary
step before the above reactions which distorts linearity of
the plot in its initial part. Model 3 describes the behavior
of the system in the presence of fibrinogen fragments.
Model 1. The dependence of tPA activity on the plasmi-
nogen concentration was investigated in the coupled reaction
assay according to the following schemes
where Pg represents plasminogen, Pn represents plasmin, K
1
and K
2
are the Michaelis constants (K
m
) of the corresponding
enzymes, and k
1
and k
2
are catalytic constants (k
cat
). The
process was followed by conversion of the substrate S-2251
(S) to the colored product (P).
The usual analysis implies transformation of the coordi-
nates ([P] versus time) and plotting of [P] versus t
2
(14).
The curves after transformation are supposed to be linear in
the initial part of the chart where concentrations of both Pg
and S may be considered as constants and are equal to their
initial values [Pg]
0
and [S]
0
. The slopes of these lines are
proportional to Michaelis equation for the first reaction which
allows one to calculate K
1
and k
1
at known K
2
and k
2
. This
method has two major limitations: (i) the linear part can be
manifested only at [S]
0
. K
2
and (ii) any prestationary
kinetics would interfere with the accuracy of determination
(like a lag phase or initial “jump”). Another model has been
developed which includes correction for the decrease in S
concentration (15).
We propose a simple kind of analysis which makes it
possible to use any convenient concentration of S and fit
tPA + Pg S
K
1
tPA-Pg
9
8
k
1
tPA + Pn (S1a)
Pn + S S
K
2
Pn-S
9
8
k
2
Pn + P (S1b)
Two Interconvertible Forms of Bovine tPA Biochemistry, Vol. 37, No. 36, 1998 12633

the curve in the whole range of [P]. Reaction schemes S1a
and S1b can be described by the corresponding system of
differential equations:
The tPA-reaction (S1a) under the chosen conditions was
slow compared to the one catalyzed by Pn (S1b). It satisfied
the requirement [Pg] ≈ [Pg]
0
(V
a
≈ const) in the working
time range and allowed a simple integration of eq 1a to [Pn]
)V
a
t. After substitution of [Pn] in eq 1b by V
a
t and
integration, the system was expressed as one equation:
A simple transformation resulted in the linear dependence
of y on t
2
where y ) 2K
2
/k
2
ln(s
0
e
p/K
2
/(s
0
- p)) and y
0
is the error in
determination of the zero point (y
0
≈ 0).
The initial coordinates p versus t were transformed to y
versus t
2
using the known values of K
2
) 250 µM and k
2
)
1000 min
-1
(16), as well as the value of s
0
from the
experiment (500 µM). The slope of an individual line was
equal to V
a
at the corresponding [Pg], and the experiments
carried out at different [Pg] gave a set of lines with different
slopes (V
a1
, V
a2
, ...), see Figure 4. The dependence of V
a
on
[Pg] can be fitted according to eq 1a in order to calculate
parameters of the tPA-catalyzed reaction (K
1
and k
1
).
The presence of any prestationary kinetics would disturb
the linearity of the chart y versus t
2
in its initial part. It
might be possible to evaluate the slope of the final linear
component (reached at the stationary conditions) disregarding
the initial shape of the curve if the prestationary step is
relatively quick. One should be careful nominating the linear
part of a curved dependence, as underestimation of the slope
can be quite dramatic due to inappropriate choice.
Model 2. A more sophisticated approach allowed us to
derive some information from prestationary kinetics as well.
From Figure 4A one can see a well defined lag phase in the
reaction with bovine tPA. Its expression increased propor-
tionally to the added Pg, and no linear component could be
found at highest [Pg] in the used time scale. This observation
FIGURE 2: (A) Zymography (lane 1) and SDS-PAGE (lane 2) of
one-chain tPA. (B) SDS-PAGE of one- and two-chain tPA.
Lanes: (1) one-chain tPA unreduced, (2) two-chain tPA unreduced,
(3) one-chain tPA reduced, (4) two-chain tPA reduced.
FIGURE 3: Inhibition of the bovine tPA-mediated plasminogen
activation by PAI-1, PAI-2, and EACA in the presence of fibrinogen
fragments. In the case of inhibition by PAI-1 and PAI-2, 2 pmol
of tPA was incubated at room temperature for 20 min with 9 pmol
PAI-1or PAI-2 before addition to the reaction mixture consisting
of 0.1 M Tris pH 7.4, 0.02% Tween 80, 0.27 µM Plg, 8.4 µg
fibrinogen fragments, and 0.5 mM S-2251 in a total volume of 0.2
mL. EACA was added to the reaction mixture just before initiation
of the reaction. The reaction velocity was calculated as the slope
of the line in a plot of the absorbance versus t
2
.
FIGURE 4: Release of p-nitroaniline after cleavage of the chro-
mogenic plasmin substrate S-2251 in the reaction medium with tPA
and Pg presented in the transformed coordinates y versus t
2
(see
eqs 3 and 6). Solid lines show the best fit according to eq 6 and
model 2. (A) The dependence was obtained for bovine tPA. The
symbols O, 4, 0, ), 3, b, and + correspond to the Pg
concentrations 0.084, 0.19, 0.30, 0.41, 0.52, 0.63, and 0.84 µM,
respectively. (B) The dependence was obtained for human tPA.
The symbols O, 4, 0, ), 3, b, and + correspond to the Pg
concentrations 0.084, 0.19, 0.30, 0.41, 0.52, 0.63, and 0.73 µM,
respectively.
d[Pn]
dt
)V
a
)
k
1
[tPA]
0
[Pg]
K
1
+ [Pg]
(1a)
dp
dt
)
k
2
[Pn](s
0
- p)
K
2
+ (s
0
- p)
(1b)
K
2
ln(s
0
/(s
0
- p)) + p )
1
/
2
k
2
V
a
t
2
(2)
y ) y
0
+V
a
t
2
(3)
12634 Biochemistry, Vol. 37, No. 36, 1998 Johnsen et al.

pointed to a longer time of the prestationary reaction at high
[Pg] when compared to that at low [Pg].
The simplest explanation of the observed phenomenon
implied the existence of two interconvertible tPA conforma-
tions being in equilibrium according to reaction scheme S2
The form tPA
*
cannot bind Pg and is referred to as the
“inactive” enzyme while the “active” tPA is involved in the
reaction with Pg, see reaction schemes S1a and S1b. A lag
phase in the dependence y versus t
2
will be visible when the
initial equilibrium tPA
*
a tPA is shifted to tPA
*
(k
-
. k
+
, [tPA]/[tPA*] , 1) and the conversion between these forms
is relatively slow. The concentration of tPA at the beginning
of the reaction was assumed to be approximately zero in
order to minimize the number of parameters in the equations.
The appearance of Pn in time depends now not only on
the velocity of the tPA reaction itself (V
a
) but also on
equilibration between tPA
*
and tPA:
The rate coefficient k
*
is responsible for the expression of a
lag phase (tPA
*
-tPA equilibration) and depends on the
plasminogen concentration. Increase in [Pg] promotes a shift
in favor of tPA + tPA-Pg which prolongs the equilibration
and decreases the rate coefficient of the prestationary phase
from k
*
) k
+
+ k
-
at [Pg] f 0tok
*
) k
+
at [Pg] f ∞. The
dependence of k
*
on [Pg] has a Michaelis-like nature with
the half saturation parameter equal to K
1
(tPA-Michaelis
constant). Another parameter in eq 4 (V
a
) is a counterpart
of V
a
in eq 1a with the exception of the affinity to
plasminogen reduced here by the factor (1 + k
-
/k
+
).
Integration of eq 4 gives a complex formula for [Pn] as a
function of time:
Substitution of [Pn] in eq 1b and integration provide the
following expression of y as a function of t:
where the notation for y and y
0
is the same as before in eq
3 and V
a
, k
*
are given in eq 4. The chart y versus t
2
has a
tendency to be linear at the sufficiently long time of the
reaction, i.e., y ≈ y
0
+V
a
t
2
at t . 1/k
*
. The curves from
Figure 4 were fitted by a nonlinear regression program using
eq 6 to calculate parameters V
a
and k
*
at different [Pg]. Their
values were plotted against [Pg] and analyzed according to
the corresponding formulas in eq 4. The relevant constants
for the tPA reaction (k
+
,k
-
,k
1
,K
1
) were estimated.
Model 3. The presence of fibrinogen fragments in the
reaction medium somewhat complicated the analysis by
adding another component to the system. The lag phase was
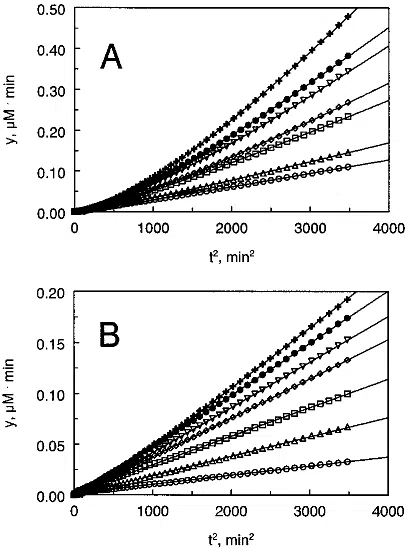
already visible both for bovine tPA and human tPA (Figure
5 A,B), and it was expressed at low [Pg] as well as at high
[Pg]. Several models, analogous to reaction scheme S2,
could have caused the appearance of the prestationary stage
with one or another formula for k
*
in eq 6. The scheme
discussed below demands some limits for the value of k
*
which makes it easy to accept or reject this model on the
basis of appropriate or inappropriate fit.
At high concentration of Fb, reaction scheme S3 is reduced
to the following description:
The above model should be supplemented to reaction
schemes S1a and S1b with Fb-tPA written instead of tPA.
The fitting can be performed on the basis of eq 6, where
k
*
) k
+
and the half-maximal value of V
a
is reached at [Pg]
) K
1
. Our attempt to apply reaction scheme S3a for human
tPA (Figure 5B) was quite successful, see the Results. On
the other hand, the fitting curves for bovine tPA (Figure 5A)
showed lower accordance with the experimental data, which
could imply existence of a more complex mechanism than
tPA* {
\
}
k
+
k
-
tPA (S2)
d[Pn]
dt
)V
a
(1 - e
-k
*
t
) (4)
where V
a
)
k
1
[tPA]
0
[Pg]
K
1
(
1 +
k
-
k
+
)
+ [Pg]
, k
*
) k
+
+
k
-
1 +
[Pg]
K
1
[Pn] )V
a
t -
V
a
k
*
+
V
a
k
*
e
-k
*
t
(5)
y ) y
0
+V
a
t
2
-
2V
a
k
*
t +
2V
a
k
*
2
(1 - e
-k
*
t
) (6)
FIGURE 5: Release of p-nitroaniline after cleavage of the chro-
mogenic plasmin substrate S-2251 in the reaction medium with tPA,
Pg, and fibrinogen fragments presented in the transformed coor-
dinates y versus t
2
(see eqs 3 and 6). Solid lines show the best fit
according to eq 6 and the model 3. (A) The dependence was
obtained for bovine tPA. The symbols O, 4, 0, ), and b correspond
to the Pg concentrations 0.05, 0.14, 0.27, 0.41, and 0.68 µM,
respectively. (B) The dependence was obtained for human tPA.
The symbols O, 4, 0, ), 3, and b correspond to the Pg
concentrations 0.05, 0.14, 0.27, 0.41, 0.54, and 0.68 µM, respec-
tively.
tPA* {
\
}
k
+
k
-
tPA + Fb S Fb-tPA (S3)
tPA*
9
8
k
+
Fb-tPA (S3a)
Two Interconvertible Forms of Bovine tPA Biochemistry, Vol. 37, No. 36, 1998 12635

![FIGURE 8: Dependence of the activity of human tPA (Va) on [Pg]. (1) The reaction was performed with 3.3× 10-4 µM tPA in the absence of fibrinogen fragments. The solid curveVa ) 0.0005Pg/ (4.5 + Pg) represents the best fit; see model 2. The dashed lineVa ) 0.0001Pg is shown as an example of an alternative fit. (2) The reaction was performed under the same conditions but in the presence of fibrinogen fragments. The solid curveVa ) 0.0085Pg/ (0.1 + Pg) represents the best fit; see model 3.](/figures/figure-8-dependence-of-the-activity-of-human-tpa-va-on-pg-1-25rq2pyu.webp)
![FIGURE 7: Dependence of the prestationary phase rate coefficient (k*) on [Pg] obtained for bovine tPA without fibrinogen fragments. The chart was approximated according to the appropriate formula in eq 4. The solid curve represents the best fitk* ) 0.3Pg/(1+ Pg/0.1)+ 0.006 which corresponds to the ratiok-/k+ ) 50; see model 2. The dashed curvek* ) 0.3Pg/(1+ Pg/0.1)+ 0.0003 is shown as an example of alternative fit which corresponds to the ratio k-/k+ ) 1000; see model 2.](/figures/figure-7-dependence-of-the-prestationary-phase-rate-2ryw3qdp.webp)