ORIGINAL PAPER
Comparative footprint of alien, agricultural and restored
vegetation on surface-active arthropods
Rembu N. Magoba
•
Michael J. Samways
Received: 21 October 2010 / Accepted: 4 April 2011
Ó Springer Science+Business Media B.V. 2011
Abstract Both invasive alien trees and agricultural
conversion have major impacts on biodiversity. We
studied here the comparative impact of these two
types of land transformation on a wide range of
surface-active arthropod species using pitfall traps,
with evergreen sclerophyllous natural vegetation
(fynbos) as the control. The study was in the Cape
Floristic Region, a global biodiversity hotspot, where
alien trees are of major concern and where vineyards
replace natural fynbos vegetation. Surface-active
arthropods were selected as they are species rich,
relatively immobile, and occur in high abundance.
We hypothesized that the impact of the two types of
land cover transformation would produce similar
qualitative and quantitative effects on the arthropods.
We also compared the results in the transformed and
natural areas with those in areas cleared of alien trees.
Arthropod species richness in cleared areas was
higher than in vineyards and more similar to that in
natural fynbos, while alien trees had the lowest.
Overall abundance scores were highest in cleared
areas, closely followed by fynbos, then vineyards and
lowest in alien trees. Several species were restricted to
each vegetation type, including alien trees. In terms of
assemblage composition, all vegetation types were
significantly different, although fynbos and vineyards
grouped, suggesting that vineyards have less impact on
the arthropod community than do alien trees. When
rare species were excluded, vineyards and cleared sites
grouped, indicating some recovery but only involving
those species that were common and habitat tolerant.
Our results suggest that vineyards retain a greater
complement of indigenous species than alien trees, but
that clearing of these aliens soon encourages estab-
lishment of indigenous species. Although there were
significant differences in soil moisture and litter depth
within and between vegetation types, we did not record
them as significantly affecting species richness or
abundance, even in alien vegetation, an encouraging
sign for restoration.
Keywords Alien vegetation Agricultural
conversion Restoration Arthropods Cape Floristic
Region
Introduction
Invasion by alien tree species is a global environ-
mental problem (Mack et al. 2000; Richardson and
Pys
ˇ
ek 2006), affecting movement patterns of animals,
including insects (Wood and Samways 1991), and
threatening their habitats (Armstrong and van
Electronic supplementary material The online version of
this article (doi:10.1007/s10530-011-9994-x) contains
supplementary material, which is available to authorized users.
R. N. Magoba M. J. Samways (&)
Department of Conservation Ecology and Entomology,
and Centre for Invasion Biology, University
of Stellenbosch, P/Bag X1, Matieland 7602, South Africa
e-mail: samways@sun.ac.za
123
Biol Invasions
DOI 10.1007/s10530-011-9994-x

Hensbergen 1996; Samways et al. 1996; Richardson
and van Wilgen 2004; Samways and Taylor 2004).
However, different ecosystems vary considerably in
their susceptibility to invasion (Chytry
´
et al. 2008),
with the impacts of alien tree species in natural
systems being dependent on invader attributes and on
characteristics of the invaded community (Mason and
French, 2008). Invasive alien trees (IATs) are wide-
spread in the Cape Floristic Region (CFR), South
Africa, and their success attributed to their good
colonizing abilities, especially in disturbed areas
(Holmes and Richardson 1999). In many parts of the
world, alien trees are the main component of
commercial forestry, with these trees often spreading
from planting sites (Simberloff et al. 2010). IATs
such as Pinus, Acacia and Eucalyptus species are of
major commercial importance in South Africa, but
they are also a threat to water supplies and biodiver-
sity (Wittenberg and Cock 2001; Le Maitre et al.
2004), especially in the CFR (Macdonald and
Richardson 1986).
Another impact on natural systems is conversion
to agriculture, which changes ecosystem composition
and function (Donald and Evans 2006), and its
biodiversity (Turin and den Boer 1988; Newton 2004;
Gaigher and Samways 2010). Agriculture is one of
the most significant human-induced disturbances that
threatens terrestrial biodiversity (Sala et al. 2000;
Tilman et al. 2001), affecting the availability of
suitable terrestrial habitats (Feber et al. 1996; Warren
et al. 1997; Jeanneret et al. 2003; Kleijn and van
Langevelde 2006). For example, vegetation structure
is important to some arthropods because it can affect
their ability to thermoregulate and reproduce (Holl
1996). However, vegetation loss for agricultural
purposes does not threaten all arthropods equally
(Fleishman et al. 1999), with some relatively small-
scale agriculture maintaining open, early seral bio-
topes favoured by some arthropod species (Shreeve
and Mason 1980; Sibatani 1980).
The regional focus for this study is the CFR, a
global biodiversity hotspot (Mittermeier et al. 2005)
and a world centre of plant diversity and endemism
(Linder 2005; Proches¸ and Cowling 2006), with 67%
of plant species endemic to the region (Linder 2005
).
It is also an important area for many rare and
endemic arthropod species (Johnson 1992; Picker and
Samways 1996; Wright and Samways 1998;
Giliomee 2003; Proches¸ and Cowling 2006; Proches¸
et al. 2009). Of significance here is that both invasive
alien trees (IATs) and agriculture, especially vine-
yards, have an impact on this biodiversity (Rouget
et al. 2003). Yet there is little knowledge on the
comparative impact, or footprint, of these two types
of human-induced land transformations on this bio-
diversity, so we investigate here the comparative
impact of IATs and vineyards on soil-surface arthro-
pod diversity, and compare it with patches where
IATs had been removed. We chose this group of
arthropods as it is species-rich, occurs in high
abundance, and most species are relatively immobile
(therefore allowing spatially-explicit interpretation of
the arthropod data). We hypothesized that the quan-
titative and qualitative adverse footprint in its entirety
(interior and edge) of IATs and vineyards on
arthropod diversity is the same, as they are both, at
least to the human eye, major transformations of
landscape matrix at the spatial scale of the patch.
Study area and methods
Study sites
Sampling was in three nature reserves and seven wine
estates within the CFR (Table 1). At each of these ten
localities, transects were established, so that the focal
land cover/land use types (hereafter referred to as
‘vegetation type’) were adjacent to each other. In
total, there were 36 transects, each 256 m in length.
Half of the transect, i.e. 128 m, was on either side of
the boundary of land use/land cover types, with the
exception of two transects (128 m in length) that
were established across native vegetation and small
IAT fragments (Table 1). Vegetation type was in four
categories: natural fynbos (evergreen schlerophyllous
shrublands characterized by graminoids of the Res-
tionaceae, and shrubs of the Ericaceae and Protea-
ceae), IATs, cleared of invasive alien trees (CIATs),
and vineyards. These resulted in six different pairs of
vegetation types (Table 1).
Natural fynbos was relatively untransformed by
human activity and selected from the nature reserve
and the wine farms with less than 10% alien tree
vegetation. Natural fynbos was predominantly moun-
tain fynbos, with common plant species being geo-
phytes Watsonia borbonica, Cyphia phyteuma and
Chasmanthe aethiopica; herbs Gymnodiscus capillaris
R. N. Magoba, M. J. Samways
123

and Dimorphotheca pluvialis; shrubs Aspalathus forb-
esii, A. aspalathoides, Lebeckia sepiaria, Lotononis
prostrata, Hymenolepis crithmoides, Protea compacta,
P. repens, P. neriifolia, and Salix species, as well
as various ericas. IATs was considered a vegetation
type with more than 90% alien trees, mainly Acacia
Table 1 Details of the study sites and transects
Category Site name Locality Pair of vegetation Transect
length (m)
No. pitfall
traps
Vineyards Vergelegen E: 34.09206
S: 18.89851
Cleared IATs Natural Fynbos 256 28
Cleared IATs Natural Fynbos 256 28
Cleared IATs Natural Fynbos 256 28
Cleared IATs Natural Fynbos 256 28
IATs Natural Fynbos 256 28
IATs Cleared IATs 256 28
IATs Cleared IATs 256 28
Bilton E:34.01431
S:18.87259
Vineyard IATs 256 28
Vineyard IATs 256 28
Vineyard IATs 256 28
Vineyard Natural Fynbos 256 28
Vineyard Natural Fynbos 256 28
Stellenzicht-Driekoppen E:34.98575
S:18.95216
Vineyard Cleared IATs 256 28
Vineyard Cleared IATs 256 28
Waterford E: 34.06625
S: 18.87626
Vineyard IATs 256 28
IATs Cleared IATs 256 28
Rustenberg E: 33.96862
S: 18.9354
Vineyard Cleared IATs 256 28
Vineyard Cleared IATs 256 28
Vineyard Natural Fynbos 256 28
Vineyard Natural Fynbos 256 28
Vineyard Natural Fynbos 256 28
Vineyard Natural Fynbos 256 28
Vineyard IATs 256 28
Dornier E: 34.01731
S: 18.86607
Vineyard IATs 256 28
Waterford-Driekoppen E: 34.0063
S: 18.87639
Vineyard Cleared IATs 256 28
Vineyard Cleared IATs 256 28
Nature Reserves Jonkershoek E: 33.98317 IATs Natural Fynbos 256 28
S: 18.94967 IATs Natural Fynbos 256 28
IATs Natural Fynbos 128 24
IATs Natural Fynbos 128 24
Helderberg E: 34.00535 IATs Cleared IATs 256 28
S: 18.8748 IATs Natural Fynbos 256 28
Hottentots Holland E: 34.06436 IATs Cleared IATs 256 28
IATs Cleared IATs 256 28
S: 18.87469 Cleared IATs Natural Fynbos 256 28
Cleared IATs Natural Fynbos 256 28
Total number of pitfall traps 1000
IATs invasive alien trees
Comparative footprint of alien, agricultural and restored vegetation
123

mearnsii, A. longifolia, A. saligna, Hakea sericea,
H. drupacea, Pinus Pinaster, P. radiata, Eucalyptus
lehmannii, E. diversicolor and Populus trees, with an
understorey of grasses and forbs. Since 2000, farmers
together with government authorities started clearing
these invasive alien trees, with follow-up clearing
undertaken at least every 3 years. Physical or manual
and chemical control methods were applied during IAT
clearing. In essence, IATs were cut and herbicides
applied to the remaining mainstem. Only organic
vineyards were considered. Organic vineyard manage-
ment involves no application of artificial fertilizers as
the soils are relatively fertile through permanent cover
crop (i.e. wheat), and only apply pesticides when
absolutely necessary. One application of chemical
agent chlorpyrifos was applied during early August to
control mealybugs which are the vectors of a viral
disease of the vines.
The 9800 ha Jonkershoek nature reserve com-
prises the Jonkershoek mountains and portions of the
upper Jonkershoek valley where large Radiata pine
(Pinus radiata) plantations are a distinctive feature
bordering fynbos vegetation. This reserve was con-
sidered due to the presence of mountain fynbos
adjacent to invasive alien trees (i.e. Pinus and Hakea
spp.). The 286 ha Helderberg nature reserve has
mountain fynbos dominated mainly by protea spe-
cies. This site was chosen for its IATs adjacent to a
site cleared of IATs (i.e. Pinus spp.). The Hottentots
Holland nature reserve is 42 000 ha and comprises
the Hottentots Holland mountains with the presence
of pristine mountain fynbos adjacent to IATs (i.e.
P. radiata).
Sampling
Sampling of the arthropods was on three occasions
(August-October 2006, May–July 2007 and Novem-
ber 2007–January 2008), when soil surface charac-
teristics (i.e. soil moisture with radioactive moisture-
density gauge (Troxler 3411-B) and leaf litter depth
by inserting a steel rod, 4 mm in diameter, into the
leaf-litter until the harder soil layer was reached)
were also measured. Pitfall traps were used to sample
arthropods as this method is particularly good for
comparative studies of soil-surface active arthropods
(Samways et al. 2010). The 256 m transects consisted
of a trap-set of two individual pitfall traps, 1 m apart,
placed at log 2 intervals: 2, 4, 8, 16, 32, 64 and 128 m
on either side of the boundary between two adjoining
vegetation types to ensure all species across
the whole vegetation type were accounted for. The
reason for this layout of traps was because the
‘footprint’ of any one patch has both interior and
edge, with a cross over at approximately 30 m
(Samways and Moore 1991; J.S. Pryke unpublished
data). The aim was to give approximate equal
weighting to both interior and edge, while at the
same time having a sampling programme that was
practical in terms of time, hence the log 2 intervals.
This is a study of a land mosaic consisting of
patchwork of land use types, making a reference site
of a small-sized patch of fynbos being more appro-
priate than an extensive nature reserve so as to
compare different types of patches of comparative
size. Furthermore, beta diversity is very high in
extensive natural habitats and we did not want to
invoke another variable into the data. It was not the
intention here to unpack all the complex details of
edge effects for the different taxa but rather to
compare patches in their entirety in a variety of
landscape contexts (see Wiens et al. 1993). However,
two transects, between IATs and fynbos, were each
four traps short, owing to unavailability of extensive
sites (Table 1). The total was 1000 pitfall traps (two
per set, fourteen sets per transect, six transects per
vegetation type pair and six vegetation pairs from
four vegetation types, minus eight traps) (Table 1).
Pitfall traps for sampling arthropods were 500 ml
plastic honey jars, each containing a replaceable
paper cups, 8 cm diameter, 12 cm deep. Each trap
was one-third filled with 70% ethylene glycol. Traps
remained closed during non-sampling periods, and
opened for five consecutive days without rain
(Borgelt and New 2006). Samples then were washed
in water, and transferred to 70% ethanol.
Sampling in vineyards was under vine rows to
minimize impact of disturbance by farm activity. All
pitfall traps were established three months prior
initial sampling to eliminate ‘digging effects’.
Arthropod samples from each trap set were com-
bined, resulting in one sample per sampling station
(i.e. 1000 pitfall traps gave 500 samples per sam-
pling period, making 1 500 samples over the three
sampling periods).
Arthropods were identified to species, or where
this was not possible, assigned to morphospecies.
Voucher specimens are in the Entomology Museum,
R. N. Magoba, M. J. Samways
123

Stellenbosch University, although spiders are in the
National Collection of Arachnida, National Museum,
Pretoria. Identification was by keys and expert
opinion.
Data analyses
Species accumulation curves, using EstimateS
version 8.0.0 with samples randomized 50 times
(Colwell 2006), were produced for all vegetation types
separately, and for all combined. Non-parametric
species estimators were used to provide the best
overall arthropod species estimates for all vegetation
types (Hortal et al. 2006). The incidence based
Coverage Estimator (ICE) is a robust and accurate
estimator of species richness (Chazdon et al. 1998),
whereas Chao2 and Jackknife estimators provide the
least biased estimates should insufficient sampling be
an issue (Colwell and Coddington 1994), and were
calculated here using EstimateS (Colwell 2006) for
all vegetation types separately and for all combined.
As arthropod species richness was normally dis-
tributed and variance homogeneous, one-way analy-
sis of variance (ANOVA) was performed on species
richness data. However, log transformations were
used for abundance data. Multiple comparisons of the
means were made using Bonferroni methodology
(Legendre and Legendre 1998). ANOVA was used to
test for differences between means of populations.
ANOVAs were performed on the selected envi-
ronmental variables in the different vegetation types
using SPSS v17 software (SPSS Inc. 2006), and
significance level was set at a P-value [0.05. Where
the ANOVAs were not significant, the analysis was
terminated. However, where the result was signifi-
cant, it was investigated further using multi-compar-
ison tests.
Classification trees, using CHAID growth limits
(SPSS Inc. 2006) determined the relationship
between the vegetation types in terms of species
richness and abundance relative to environmental
variables. Significance level for splitting nodes and
merging categories was 0.05, and the significance
values were adjusted using Bonferroni methodology.
Correlations between species richness and abundance
with environmental variables for all vegetation types,
separately and collectively, were calculated using
Spearman’s Rank Order Coefficient, since the data
were not normally distributed.
Multivariate analysis, using Primer Ver. 5 (Clarke
and Gorley 2001), was used to detect trends and to
explore the differences in arthropod assemblages
between different vegetation types. Bray-Curtis sim-
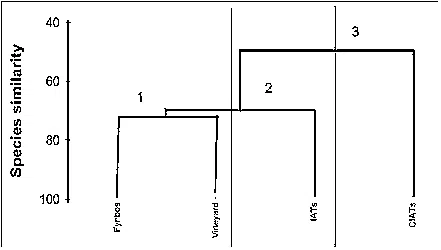
ilarity coefficients were used to derive similarity
matrices of arthropod data, then clustering dendro-
grams and non-metric, multi-dimensional scaling
(NMDS) ordination plots to detect trends in similar-
ity. The main advantage of NMDS is its greater
ability to represent complex relations accurately in
low-dimensional space (Clarke and Warwick 2001).
Joint absences in the NMDS were ignored to
emphasize similarity in common or rare species,
comparing only percentage composition (Clarke and
Warwick 2001). The dendrograms were group-aver-
age linking on Bray-Curtis species similarities from
standardized abundance data.
Principal Components Analysis (PCA) was used to
determine whether arthropods from the different
vegetation types fell into distinct groups (Clarke
and Warwick 2001). PCA considers a different
starting point by making different assumptions about
the definition of (dis)similarity of samples being
compared. Rare species were excluded from the PCA,
so that the species retained were more comparable
with the number of samples (Clarke and Warwick
2001). Distances between vegetation types on the
ordination attempt to match the corresponding dis-
similarities in arthropod assemblage composition i.e.
similar vegetation types would have very similar
arthropod assemblages, while vegetation types that
are dissimilar would have few species in common, or
the same species at very different levels of abun-
dance. However, PCA can produce inconclusive
results, so the data were further analyzed using
cluster analysis for determining how the vegetation
types varied, and how they grouped in terms of their
arthropod assemblages. Cluster analysis produces
results that broadly agree with PCA (Clarke and
Warwick 2001), and usually it is much easier to
observe grouping in cluster analysis than in PCA.
Nevertheless, it may be easier to understand what the
groupings indicate if they have been produced by
PCA. Therefore, using a combination of cluster
analysis and PCA is an effective approach.
Analysis of similarity (ANOSIM) tests the hypoth-
esis that there are no assemblage differences between
groups of samples specified, here vegetation type
(Clarke and Warwick 2001). R is approximately zero
Comparative footprint of alien, agricultural and restored vegetation
123