





page -1-
COMPENSATORY GROWTH IN DOUBLE MUSCLED BULLS
1
2
3
Different Periods of Feed Restriction Before Compensatory Growth in Belgian Blue Bulls:
4
I. Animal Performance, Nitrogen Balance, Meat Characteristics and Fat Composition
1
5
6
J. L. Hornick
2
, C. Van Eenaeme, A. Clinquart, M. Diez, and L. Istasse
7
8
9
10
Department of Nutrition, Veterinary Faculty, Sart Tilman
11
B43 4000 Liège, Belgium
12
13
Phone: 32-(0)4-3664139
14
Fax: 32-(0)4-3664122
15
E-mail: HORNICK@.STAT.ULG.FMV.AC.BE.
16
17
18
19
20
21
1
The IRSIA (Institut pour l'Encouragement de la Recherche dans l'Industrie et l'Agriculture,
Brussels, Belgium) is gratefully acknowledged for financial help.
2
To whom correspondence should be addressed.

page -2-
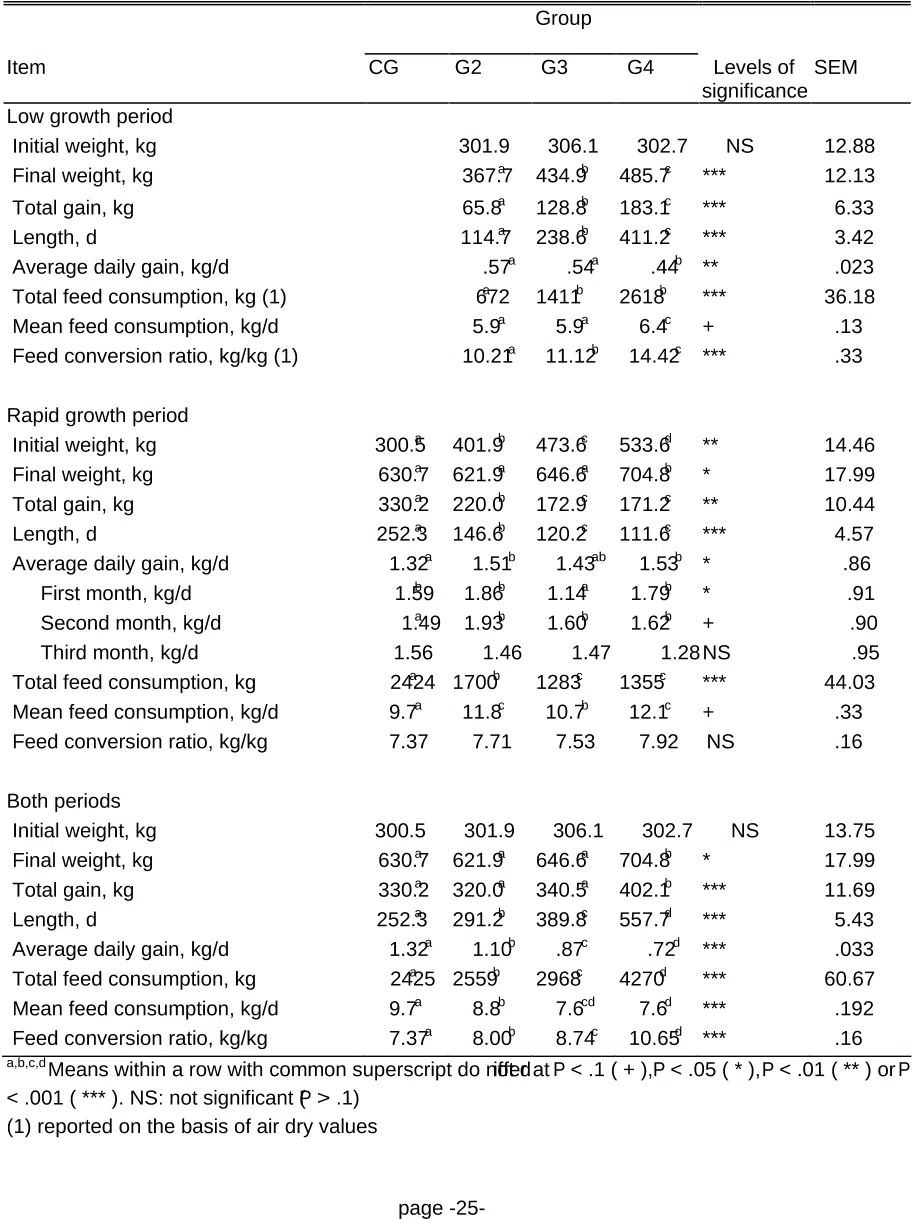
ABSTRACT: Thirty double-muscled Belgian Blue bulls were maintained at a rate of gain of .5 kg/d
1
during four length of time, 4 (G2), 8 (G3) or 14 (G4) mo (low growth period, LGP), before fattening
2
(rapid growth period, RGP). Ten control animals (CG) were fed a high-energy, high-protein diet. The
3
G2, G3, and G4 were fed a low-energy, low-protein diet during LGP and the same diet as CG during
4
RGP. Live weight was recorded biweekly, feed consumption (FC) daily, and nitrogen balance at 3
5
occasions in each group. At the slaughterhouse, the 7, 8, and 9th ribs were removed to determine
6
carcass composition, meat quality, and meat and fat composition. Compensatory growth reached a
7
maximum 2 mo after refeeding. The G2 and G4 exhibited compensatory growth ( P < .05) and had
8
higher daily FC ( P < .001). Feed conversion ratio (FCR) increased sharply after refeeding. Nitrogen
9
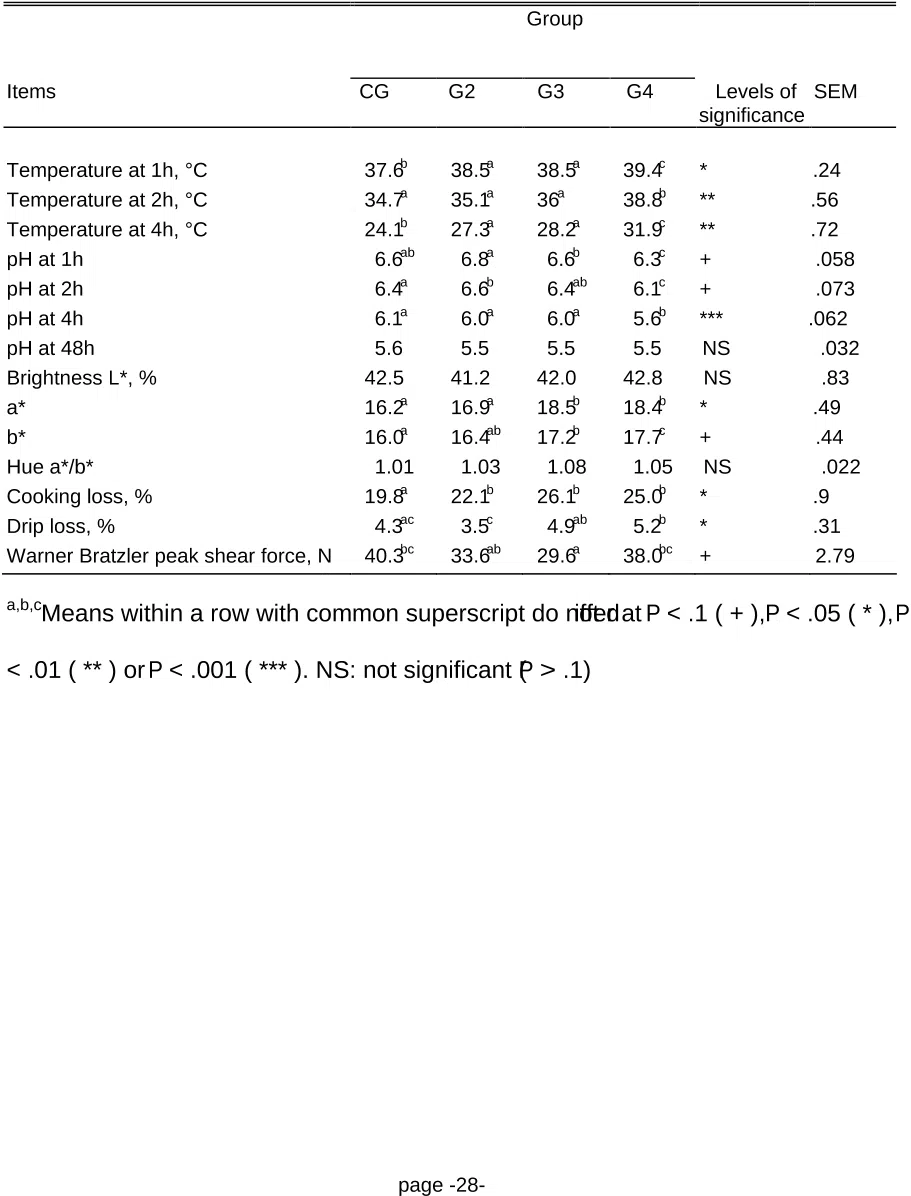
balance was higher in compensating groups ( P < .05). Compensating animals had higher carcass
10
connective and adipose tissue contents ( P < .05) but lower meat fat content (P < .05). Cattle exhibiting
11
compensatory growth had higher redness, yellowness, hue, cooking losses and drip losses, but tended
12
to have lower Warner-Bratzler peak shear force (WBPSF) values. The saturated fatty acid (SFA)
13
content of the fat decreased with the length of the LGP. Compensatory growth in double-muscled bulls
14
at the expense of higher feed intake increased peripheral fat but decreased intramuscular fat deposition.
15
16
Key Words: Belgian Blue Bulls, Compensatory Growth, Animal Performance, Carcass, Meat, Fatty
17
Acids
18

page -3-
Introduction
1
2
Compensatory growth is the ability of an animal to exhibit, after disease (Thomas et al., 1978) or
3
feed restriction (Wilson and Osborn, 1960), larger growth rates than in unaffected animals of the same
4
chronological age. In cattle, compensatory growth is well expressed when feed restriction occurs at a
5
relatively late stage of life (Berge, 1991; Berge et al., 1991). Factors contributing to compensation are
6
increases in feed intake (Baker et al., 1992), increases in gut-fill weight, or higher efficiency of feed
7
utilization (Carstens et al., 1991). The response varies according to the pattern of undernutrition and
8
realimentation, and stage of development of the animal (Wilson and Osborne, 1960). The Belgian Blue
9
breed, double-muscled type, is a large beef breed with early maturity, characterized by high average
10
daily gain, low feed conversion ratio, and high quality of carcass (Clinquart et al., 1991). Currently,
11
there is no published work on compensatory growth in Belgian Blue bulls. Therefore, an experiment
12
was conducted with Belgian Blue bulls in order to study the effects of a restricted growth, lasting for
13
three different durations, on fattening performances. Results are presented in 2 papers. This paper
14
summarizes animal performance, nitrogen balance, and carcass, meat, and fat characteristics.
15
16
Materials and Methods
17
18
Animals and Management
19
The Animal Care and Use Council of our institute approved the use and treatment of animals in
20
this study. A total of 40 Belgian Blue bulls, double-muscled type, initial age and weight range of 9.7
21
mo and 310 ± 38 kg, were divided into four groups of similar live weight. In each group, four animals
22
were randomly penned in individual stalls allowing for collection of urine and feces, and the remaining
23

page -4-
six were housed in a stanchion barn with straw as bedding. Each group was randomly assigned to one
1
of the four treatments. The first group (control, CG) was given from the beginning ad libitum access to
2
a fattening diet allowing for rapid growth. The fattening diet was based on sugar beet pulp
3
complemented with cereals, protein from vegetable origin, and a mineral mixture (Table 1). During
4
three periods with different lengths of time, the other groups received a limited quantity of a low-
5
energy, low-protein diet calculated to support an ADG of .5 kg daily gain (LGP, low growth period).
6
The low growth diet was based on pelleted straw complemented with dried lucerne, cereals, protein
7
from vegetable origin, and mineral mixture. The three groups, namely groups 2, 3, and 4 (G2, G3, G4),
8
received the low-growth diet for 115, 239, and 411 d, respectively. Subsequently, G2, G3 and G4 were
9
adapted to the concentrate fattening diet over a 15-d period of transition. The amount of concentrate
10
feed was then progressively increased and animals were allowed to consume their ration on an ad
11
libitum basis for about 1 mo after the beginning of the transition period. The concentrate diet was
12
offered during the rapid growth period (RGP) which lasted until the animals were slaughtered. The
13
animals were fed twice daily at 0600 and 1400 and were slaughtered per group when mean live weight
14
reached at least 600 kg and when the average daily gain (ADG) was lower than 1 kg/d at two
15
consecutive measurements.
16
17
Measurements
18
Feed intake of the bulls was recorded each day and live weight at 15-d intervals. Feed samples
19
were withdrawn at regular intervals for chemical analysis. At the slaughterhouse, abdominal fat was
20
removed from the carcass. Carcass weight was recorded and pH of both Longissimus thoracis muscles
21
were measured (7, 8, 9 ribs) 1, 2, and 4 h postmortem using a Portamess 751 knick pH-meter (Knick
22

page -5-
GmbH & Co, Berlin, Germany) with an Ingold "penetration" pH-electrode (Ingold AG, Urdorf,
1
Switzerland).
2
Two days after slaughter, the 7, 8, and 9th ribs were removed from the carcass. They were
3
dissected in order to separate lean meat, fat and connective tissue, and bones. Regressions of Martin
4
and Torreele (1962) for double muscled cattle were then used to assess the composition of the carcass.
5
Meat quality was determined from one 2.5-cm-thick cut of the longissimus thoracis muscle. Five
6
measurements of the final pH were performed on this cut at 48 h postmortem using the technique
7
described above. At the same time, the HunterLab Labscan II device was used for objectively
8
measuring CIE Lab brightness (L*), redness (a*) and yellowness (b*) on 5 spots 2.5 cm diameter. Hue
9
was estimated by a*/b* ratio. Seven days later, the cut was weighed in order to estimate drip loss, and
10
heated in open plastic bags in a waterbath for 50 min at 75°C. After heating, they were cooled in cold
11
tap water to room temperature, bags were drained, and cuts were mopped gently dry with paper tissue.
12
The difference between raw and heated weights was recorded as cooking loss and expressed as a
13
percentage of the raw weight. Warner-Bratzler shear force was determined with a Lloyd LR5K
14
perpendicular to the fiber direction on 10, 1.25-cm-diameter cores obtained from the heated cuts.
15
The dry matter, ash, ether extract, and crude protein concentrations of the diets were determined
16
according to official procedures (AOAC, 1975). The lipids from peripheral, intermuscular and
17
intramuscular fat samples were extracted and saponified as described by Ter Meulen et al. (1975). The
18
fatty acid composition of fat samples was determined by gas chromatography.
19
20
Statistical Analysis and Mathematical Modelling
21
Bulls were blocked (n = 10) by group (Figure 1). One-way analysis of variance, using group as a
22
factor of variation, was used to analyze data. Data relative to muscle, connective and adipose tissue,
23