![Figure 10. Stability diagram of a multi-drop system for varying forward rate constant k and drop radius R. A steady state exist within the continuous curve. Outside this region all drops dissolve (downward arrows). Outside the grey region, drops are unstable against Ostwald ripening and coarsen (upward arrows). In the grey region drops are stable against Ostwald ripening. The stability-instability boundary is shown by a dashed curve. The analytical results for the critical radii in the small drop regime are showed by the upper dotted line (Ru, Eq. (14)) and lower dotted line (Rl). The analytical results for the critical rates kc, kl and ku (Eqs (15), (20) and (21)) are showed by arrows. Large drops dissolve beyond ku, all drops dissolve beyond kc, and large drops are stable against Ostwald ripening beyond kl. The large drop regime is coloured in blue and the boundary between the large and the small drop regime is R = ξ (Eq. (11)). Parameters: h = 10−2s−1, lc = 10−2µm, D = 1µm2s−1, P̂in = 10 −1ν−1, P̂out = 10−4ν−1, φ = 5 × 10−4ν−1, where ν is the molecular volume of P and S and can be chosen arbitrarily. Reprinted figure with permission from [Wurtz J.D. and Lee C.F., Physical Review Letters, vol. 120, 078102, 2018]. Copyright (2018) by the American Physical Society.](/figures/figure-10-stability-diagram-of-a-multi-drop-system-for-1j3kapn7.webp)
![Figure 15. Our discussion in Sect. 3.4 demonstrates that in two spatial dimensions, the large distance properties of an incompressible active fluid in the ordered phase, e.g., incompressible flock, has the same scale-invariant properties as a two-dimensional smectic liquid crystals and the KPZ surface growth model. The findings thus suggest that instead of the apparent equivalence between an incompressible flock and a two-dimensional lattice depicted by the landscape in Escher’s Day and Nights [top figure (awaiting approval of use from the copyright holder)], the flock should instead be mapped onto a smectics-like layered structure such as sedimentary rocks [66] [Top half of bottom figure: photograph by Tommy Hansen. Public domain. Bottom half of bottom figure: photograph by Ryan McGrady, distributed under a CC-BY 4.0 license.].](/figures/figure-15-our-discussion-in-sect-3-4-demonstrates-that-in-2fdxeus6.webp)

![Figure 6. Monte Carlo simulation of phase separation in a ternary mixtures with non-equilibrium chemical reactions [29]. Molecules P (red dots) form energetic bounds with neighbouring P , while molecules S (blue dots) and C (not shown) do not form bounds. Chemical reactions transform P into S and vice versa with fixed reaction rate constants (Eq. (6)). In the initial state molecules are randomly distributed (leftmost snapshot). As time progresses drops form and undergo Ostwald ripening leading to increase of the average drops radius (middle snapshot). Eventually a multi-drop steady state is reached (rightmost snapshot). At the steady state drops have roughly the same radius and are evenly distributed. Simulations details can be found in [29]. Reprinted figure with permission from [Wurtz J.D. and Lee C.F., Physical Review Letters, vol. 120, 078102, 2018], Copyright (2018) by the American Physical Society](/figures/figure-6-monte-carlo-simulation-of-phase-separation-in-a-2vzv525w.webp)

![Figure 1. Cytoplasmic phase separation and tissue dynamics as active matter. a) In many distinct types of cells, certain proteins can phase separate from the cytosol to assemble membrane-free organelles, such as the stress granules (yellow drops) shown here in human epithelial cells (HeLa) [6], akin to oil drop formation in an oil-water mixture (b). c) In a monolayer of Madin-Darby Canine Kidney (MDCK) cells, the cells in the tissue can undergo dynamical rearrangement as shown by the snapshot of the velocity field shown in (d) [7]. Figure a) is adapted from [J.R. Wheeler et al., eLife vol. 5, pp. e18413, 2018], licensed under CC BY 4.0. Figure b): c©Nikola Bilic, Dreamstime.com. Figures c) and d) reprinted from Biophysical Journal, vol. 98, Petitjean et al., Velocity fields in a collectively migrating epithelium, pp. 1790-1800, Copyright (2010), with permission from Elsevier.](/figures/figure-1-cytoplasmic-phase-separation-and-tissue-dynamics-as-cn0xofov.webp)

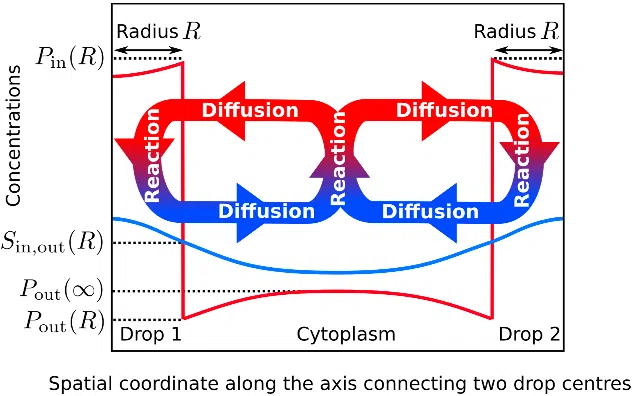
![Figure 4. Model of non-equilibrium phase separation in the cell cytoplasm. We consider the ternary mixture from Sec. 2.2 with the addition of chemical reactions. Molecules P can convert into S at rate k and vice versa at rate h (Eq. (6)). S do not participate to phase separation i.e. its concentration remains homogeneous (Ŝin = Ŝout). Figure reprinted from [Wurtz J.D. and Lee C.F., New Journal of Physics, vol. 20, no. 4, 045008, 2018], licensed under CC BY 3.0.](/figures/figure-4-model-of-non-equilibrium-phase-separation-in-the-16k650kt.webp)

![Figure 13. The flow of the two coupling coefficients g (b) ` and g (λ) ` under a DRG transformation for d = 3 to order . There are one stable and two unstable fixed points. The stable fixed point (red square) corresponds to the incompressible active fluid universality class found recently [57]. The unstable fixed points lying on the g(b)-axis (purple circle) corresponds to the ferromagnets with dipolar interactions universality class found in 1973 [58]; while the unstable fixed point lying on the g(λ)-axis corresponds to the randomly stirred fluid universality class found in 1977 (model B in [41]). Figure reprinted from [Chen L., J. Toner and Lee C.F., New Journal of Physics, vol. 17, 042002, 2018], licensed under CC BY 3.0.](/figures/figure-13-the-flow-of-the-two-coupling-coefficients-g-b-and-31zvvndn.webp)
![Figure 5. Non-driven and driven chemical reactions in phase separating systems. We consider here for clarity a particular example from our ternary mixture, where P molecules (red disks) form bonds (black lines) with neighbouring P molecules while molecules S (blue disks) do not interact. The chemical conversion of a given P into a S requires an activation energy ∆U to break these bounds. Since drops are enriched in P , many bonds need to be broken and ∆U is high (upper graph). Conversely the cytoplasm is poor in P and ∆U is thereby small (lower graph). At thermal equilibrium the chemical conversions are non-driven and energy required to overcome the barrier ∆U is provided by thermal fluctuations alone, and therefore the reaction rate constant k decreases exponentially with ∆U [31]. As a result, k is thus concentration-dependent: k is small inside drops where ∆U is high, and large in the cytoplasm. In the case of driven chemical reactions, such as ATP-dependent phosphorylation, an external source of energy is provided [18] (yellow arrows). Hence k depends on the specificities of the reaction and can potentially be independent of ∆U i.e. concentration-independent. The rightmost part of the figure is adapted from [J.R. Wheeler et al., eLife vol. 5, e18413, 2018], licensed under CC BY 4.0.](/figures/figure-5-non-driven-and-driven-chemical-reactions-in-phase-qi7h0idr.webp)




Novel physics arising from phase transitions in
biology
Chiu Fan Lee
Department of Bioengineering, Imperial College London, South Kensington
Campus, London SW7 2AZ, U.K.
E-mail: c.lee@imperial.ac.uk
Jean David Wurtz
Department of Bioengineering, Imperial College London, South Kensington
Campus, London SW7 2AZ, U.K.
Page 1 of 38 AUTHOR SUBMITTED MANUSCRIPT - JPhysD-117020
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60

Novel physics arising from phase transitions in biology 2
Abstract. Phase transitions, such as the freezing of water and the
magnetisation of a ferromagnet upon lowering the ambient temperature, are
familiar physical phenomena. Interestingly, such a collective change of behaviour
at a phase transition is also of importance to living systems. From cytoplasmic
organisation inside a cell to the collective migration of cell tissue during
organismal development and wound healing, phase transitions have emerged
as key mechanisms underlying many crucial biological processes. However, a
living system is fundamentally different from a thermal system, with driven
chemical reactions (e.g., metabolism) and motility being two hallmarks of its non-
equilibrium nature. In this review, we will discuss how driven chemical reactions
can arrest universal coarsening kinetics expected from thermal phase separation,
and how motility leads to the emergence of a novel universality class when the
rotational symmetry is spontaneously broken in an incompressible fluid.
1. Introduction
Collective phenomena are intimately linked to the phenomenon of phase transitions
in physics. At a typical phase transition, a many-body system with constituents that
interact only locally with their neighbours, be they molecules or living organisms,
can collectively change their behaviour upon a subtle change of a single parameter,
to the extent that the qualitative behaviour of the whole system is modified. Phase
transitions encompass many everyday phenomena such as oil drop formation in a
salad dressing and magnetisation in some metals. The study of phase transitions is of
fundamental interest to physicists because of the emergence of universal behaviours
at a phase transition. By a universal behaviour, we mean certain properties of the
system that are highly independent of the system’s microscopic details. In the salad
dressing example, such property can be the power law exponent that governs how the
average size of oil drops changes with time; in the example of magnetisation, it can be
the power law exponent that governs how the correlation function of two atomic spins
decays with respect to their distance. Recently, phase transitions in living systems
have also been under intense attention. Indeed, the generic non-equilibrium nature
of biological systems have given rise to novel universal behaviours not seen before.
In this review, we will focus on two such examples: phase separation with driven
chemical reactions, motivated by the mechanism underlying the formation of some
non-membrane bound organelles in cells [1, 2], and spontaneous symmetry breaking
in incompressible active matter, motivated by its relevance to biological tissues [3, 4, 5]
(Fig. 1).
In Sect. 2, we will first describe the relevance of phase separation in cytoplasmic
organisation and then review the latest findings on how driven chemical reactions
(e.g., adenosine triphosphate (ATP)-driven phosphorylation) can lead to co-existing
phase-separated protein drops in the cytoplasm, contrary to the universal coarsening
behaviour expected in its equilibrium counter part. In Sect. 3, motivated by the
collective behaviour found in motile organisms, we will introduce a generic model of
incompressible active fluids from a symmetry consideration. We will then elucidate
how a novel critical behaviour emerges at the onset of collective motion, and discuss
the universal behaviour of a two dimensional incompressible active fluid in the ordered
phase. Finally, we will end with Conclusion & Outlook.
Page 2 of 38AUTHOR SUBMITTED MANUSCRIPT - JPhysD-117020
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60

Novel physics arising from phase transitions in biology 3
Figure 1. Cytoplasmic phase separation and tissue dynamics as active matter.
a) In many distinct types of cells, certain proteins can phase separate from the
cytosol to assemble membrane-free organelles, such as the stress granules (yellow
drops) shown here in human epithelial cells (HeLa) [6], akin to oil drop formation
in an oil-water mixture (b). c) In a monolayer of Madin-Darby Canine Kidney
(MDCK) cells, the cells in the tissue can undergo dynamical rearrangement as
shown by the snapshot of the velocity field shown in (d) [7]. Figure a) is adapted
from [J.R. Wheeler et al., eLife vol. 5, pp. e18413, 2018], licensed under CC
BY 4.0. Figure b):
c
Nikola Bilic, Dreamstime.com. Figures c) and d) reprinted
from Biophysical Journal, vol. 98, Petitjean et al., Velocity fields in a collectively
migrating epithelium, pp. 1790-1800, Copyright (2010), with permission from
Elsevier.
2. Non-equilibrium phase separation: a mechanism for cytoplasmic
organisation
2.1. Membrane-less organelles
Biological cells organise their contents in distinct compartments called organelles,
typically enclosed by a lipid membrane that forms a physical barrier and controls
molecular exchanges with the surrounding cytosol. Recently an intriguing class
of organelles lacking a membrane is being studied intensely [8]. Membrane-less
organelles have attracted an intense interest from the biology community as they
are present in many organisms from yeast to mammal cells, and are critical for
multiple biological functions. For example P granules are involved in the asymmetric
division of the Caenorhabditis elegans embryo [9], and stress granules assemble during
environmental stress and protect cytoplasmic RNA from degradation [10] (Fig. 1
a)). Membrane-less organelles are generally spherical, fuse together upon contact
[11, 12], and their components quickly shuttle in and out [13, 14], thus resembling
liquid drops. Indeed, strong experimental evidence indicates that membrane-less
Page 3 of 38 AUTHOR SUBMITTED MANUSCRIPT - JPhysD-117020
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60

Novel physics arising from phase transitions in biology 4
organelles are assembled via liquid-liquid phaseLiving and engineered systems rely
on the stable coexistence of two interspersed liquid phases. Yet, surface tension
drives their complete separation. Here, we show that stable drops of uniform and
tunable size can be produced through arrested phase separation in an elastic matrix.
Starting with a cross-linked, elastic polymer network swollen by a solvent mixture,
we change the temperature or composition to drive demixing. Droplets nucleate and
grow to a stable size that is tunable by the network cross-linking density, the cooling
rate, and the composition of the solvent mixture. We discuss thermodynamic and
mechanical constraints on the process. In particular, we show that the threshold for
macroscopic phase separation is altered by the elasticity of the polymer network, and
we highlight the role of correlations between nuclei positions in determining the drop
size and polydispersity. This phenomenon has potential applications ranging from
colloid synthesis and structural color to phase separation in biological cells. separation
[2, 15, 16], a common phenomenon in every day life responsible for example for oil
drop formation in water (Fig. 1 b)). Under the equilibrium condition phase separation
is well understood [17]. However cells are driven away from equilibrium by multiple
energy-consuming processes such as ATP-driven protein phosphorylation [18], which
can potentially affect the phase-separating behavior of membrane-less constituents.
For example P granules do not distribute homogeneously in the cytoplasm but
preferentially to the posterior side of the cell [19], and stress granules form and dissolve
according to environmental cues [20]. The fascinating physics associated to membrane-
less organelles are only beginning to be investigated [12, 21, 19, 11, 22, 23, 24].
In this section, we will start with a brief summary of relevant principles of
equilibrium phase separation in Sec. 2.2. We will then review the latest progress
on phase separation driven out of equilibrium by energy-driven chemical reactions in
Sec. 2.3. Specifically we will focus on a ternary fluid model of the cell cytoplasm where
chemical reactions can convert phase-separating molecules into soluble molecules and
vice versa. We will show how such reactions can control drops assembly and size,
and suppress Ostwald ripening, allowing a collection of organelles to coexist in the
cytoplasm.
2.2. Equilibrium phase separation
Interactions between molecules can cause a homogeneous system to undergo a phase
separation, i.e. the spontaneous partitioning of a system into multiple phases of
distinct properties such as concentration [17]. The transition from the homogeneous
state to the phase-separated state is controlled by parameters such as temperature,
pressure or concentrations. The set of parameters leading to phase separation are
represented in a phase diagram as shown in Fig. 2, for a ternary mixture composed
of molecules P (red disks), S (blue disks) and C (not shown). The molecular
concentrations are labelled by the same symbols P, S, C. We assume incompressibility
and that all three types of molecules occupy the same volume, so the combined
concentration ψ ≡ P + S + C is homogeneous. The concentration C at any
point in the phase diagram is therefore given by ψ − P − S. Outside the phase
boundary (green curve) the system is homogeneous (“” symbol). Inside the phase
boundary (“♦” symbol) the system phase separates into two phases (“in” and “out”)
of distinct concentrations (
ˆ
P
in,out
,
ˆ
S
in,out
), given by the intersections between the tie-
lines (straight lines) and the phase boundary.
At the equilibrium condition a multi-drop system is unstable due to Ostwald
Page 4 of 38AUTHOR SUBMITTED MANUSCRIPT - JPhysD-117020
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60

Novel physics arising from phase transitions in biology 5
Figure 2. Equilibrium phase diagram of a ternary mixture composed of
molecules P (red discs), S (blue discs) and C (not shown). Outside the phase
boundary (green line) the system is homogeneous (“” symbol). Inside the phase
boundary (“♦” symbol) the system phase separates into two phases “in” and
“out” of distinct concentrations. The coexistence concentrations
ˆ
P
in,out
,
ˆ
S
in,out
are given by the intersections between the tie-lines (straight lines) and the phase
boundary.
ripening that causes large drops to grow and small drops to evaporate [25, 26], and
coalescence caused by the fusion of drops upon contact [27]. Eventually a unique drop
remains in a finite system. Since the crowded environment of the cytoplasm inhibits
the diffusion of macromolecular aggregates [28] we will ignore drop coalescence in this
review and focus on Ostwald ripening.
Ostwald ripening is caused by two ingredients. One is the Gibbs-Thomson relation
that relates the coexistence concentration to the drop radius. For example for the P
concentration we have:
P
in
(R) =
ˆ
P
in
(1)
P
out
(R) =
ˆ
P
out
1 −
ˆ
P
out
l
c
R
!
, (2)
were l
c
is a capillary length and
ˆ
P
in,out
are the coexistence concentrations for a flat
interface (R → ∞, Fig. 2). The smaller the drop, the larger the concentration outside
which is a consequence of the Laplace pressure [17].
The second ingredient driving Ostwald ripening is the existence of a diffusive
concentration profile between drops, which can be approximated by an ideal gas
diffusion profile in the case of small concentration outside drops [26]:
∂P
out
(r, t)
∂t
= D∇
2
P
out
(r, t) , (3)
Page 5 of 38 AUTHOR SUBMITTED MANUSCRIPT - JPhysD-117020
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60